














چگونگی عمل داروها
پيش از آنکه داروهاي سولفا در سال ۱۹۳۵ کشف شوند ، اطلاعات پزشکي داروها محدود بود . در آن زمان ، احتمالاً فقط دوازده دارو يا بيشتر داراي ارزش پزشکي مشخص بودند . بيشتر آنها عصاره هاي گياهان بودند ( مثل ديژيتاليس که از گل انگشتانه استخراج مي شود ) در حاليکه داروهايي چون آسپيرين رابطه شيميايي نزديکي با عصاره هاي گياهي دارند . ( در اين موارد ، اسيدساليسيليک را مي توان مثال زد که از درخت بيد استخراج مي گردد . )
پزشک امروز بسيار بهتر از پزشک ديروز از پيچيدگي کار دارو در بدن و محدوده اثرات آن بر بدن چه از نظر فوائد و چه از نظر عوارض نامطلوب آگاهي دارد . وي همچنين مي تواند تشخيص دهد که بعضي داروها بطور خطرناکي با داروهاي ديگر يا بعضي غذاها تداخل مي کنند .
* عمل داروها
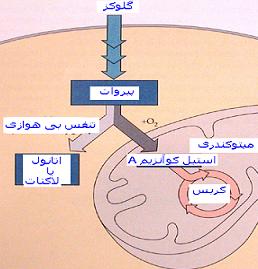
اگر چه عمل بعضي از داروها بطورکامل شناخته نشده است اما دانش پزشکي بسياري از وظايف دارو را بهنگام ورود در بدن شناسايي کرده است . البته داروها وظايف گوناگوني انجام مي دهند ، گاهي اوقات علاج بيماري مي کنند و گاهي فقط نشانه هاي بيماري را التيام مي دهند و اثر آنها در بخشهاي مختلف بدن ظاهر مي شود . هرچند که داروهاي گوناگون به روشهاي مختلف عمل مي کنند اما عمل آنها بطور کلي به سه دسته تقسيم مي شود .
* جايگزين کردن مواد شيميايي از دست رفته
بدن براي آنکه بتواند خوب کار کند بايد برخي مواد شيميايي را به مقدار کافي داشته باشد ، مثل ويتامينها و مواد معدني که بدن از غذا بدست مي آورد . يک غذاي متعادل معمولاً مواد لازم را به بدن مي دهد . اما در صورت بروز کمبودها بيماريهاي مختلفي پديد مي آيند . مثلاً فقدان ويتامين C موجب اسکوروي ، فقدان ويتامين D موجب راشيتيسم و فقدان آهن موجب کم خوني مي شوند .
از اين جمله بيماريها ، بيماريهاي ناشي از فقدان هورمونها هستند . هورمونها موادي هستند شيميايي که توسط غدد توليد مي شوند و به صورت « پيام آوران » داخلي بدن عمل مي کنند . ديابت شيرين ، بيماري آديسون و کم کاري تيروئيد همگي از کمبود هورمونهاي گوناگون بوجود مي آيند .
بيماريهائي که در اثر کمبود مواد ايجاد مي شوند با داروهايي معالجه مي شوند که مي توانند فقدان مواد مورد نياز را جبران کنند و در صورت کمبود هورمون ، جايگزينهاي حيواني يا مصنوعي مي توانند اين عوارض را برطرف کنند .
* مداخله در کار سلول
بسياري از داروها مي توانند با تحريک يا کاهش مقدار طبيعي فعاليت سلولها طرز کار سلولها را تغيير دهند . به عنوان مثال ، التهاب به علت اثر برخي از هورمونهاي طبيعي يا مواد شيميايي بر روي عروق خوني و سلولهاي خوني ديگر است . داروهاي ضد التهاب جلوي اين عمل هورمون را مي گيرند و يا توليد آنها را کند مي کنند . داروهايي که به روش مشابه عمل مي کنند براي درمان برخي بيماريها مثل اختلالات هورموني ، مشکلات انعقاد خون و بيماريهاي قلب و کليه استفاده مي شوند .
يک پيام مثل انقباض عضله از مغز ارسال شده و از طريق پايانه گيرنده آن وارد سلول عصبي مي شود . اين پيام که به صورت تکانه الکتريکي است ، عصب را طي مي کند . در پايانه يک ماده شيميايي موسوم به « ناقل عصبي » رها مي شود و پيام را از شکافي که آنرا از يک سلول عصبي مجاور جدا مي کند هدايت مي کند . اين جريان تکرار مي شود تا زمانيکه پيام به عضله مناسب برسد .
بسياري از داروها مي توانند اين جريان را تغيير دهند واين کار را اغلب با تأثير بر محلهاي گيرنده در سلولها انجام مي دهند . برخي از داروها ( آگونيست يا سازگارها ) فعاليت سلولي را تشديد و برخي ديگر ( آنتاگونيستها يا ناسازگارها ) فعاليت آنرا کم مي کنند .
فعاليت عليه ارگانيسمهاي مهاجم يا سلولهاي غيرطبيعي عامل بيماريهاي عفوني که عبارتند از ويروسها ، باکتريها ، پروتوزواها و قارچهاي مهاجم به بدن نيز يک مورد ديگر از اعمال اين داروها است . در حال حاضر ما داروهاي بسيار گوناگوني براي نابودي اين موجودات در اختيار داريم ، داروهايي که مي توانند جلوي تکثير آنها را بگيرند و يا مستقيماً آنها را از بين ببرند . داروهايي نيز هستند که با کشتن سلولهاي غيرطبيعي ، مثل سلولهاي سرطاني ، بيماري را معالجه مي کنند .
* محل هاي گيرنده
تصور مي رود بسياري از داروها اثرات خود را بر محلهاي خاصي موسوم به گيرنده هاي واقع در سطح سلولهاي بدن اعمال مي کنند . مواد شيميايي طبيعي بدن مثل ناقلهاي عصبي به اين محلها مي چسبند و پاسخي در سلول ايجاد مي کنند . ممکن است سلولها گيرنده هاي فراواني داشته باشند که هر کدام از آنها به مواد شيميايي مختلفي در بدن ميل ترکيبي دارند . داروها نير مي توانند با اتصال به گيرنده ها يا باعث افزايش اثر مواد شيميايي طبيعي بدن و پاسخ سلولي ( داروهاي آگونيست ) شوند و يا باعث متوقف کردن پاسخ سلولي ( داروهاي آنتاگونيست ) شوند .
اثرات داروها
قبل از اينکه پزشک براي درمان يک شخص بيمار اقدام به انتخاب دارو کند بايد دقيقاً فوايد و مضرات آن دارو را سبک سنگين کند . مسلماً پزشک نتيجه مثبتي از دارو انتظار دارد و مي خواهد بيماري را معالجه کند يا دست کم نشانه هاي بيماري را برطرف نمايد . اما در عين حال بايد به خطرات آن نيز توجه کند زيرا همه داروها مي توانند مضر باشند و ضرر بعضي از داروها بسيار بيشتر از بعضي داروهاي ديگر است .
* زمان واکنش
بعضي از داروها مي توانند خيلي سريع نشانه هاي بيماري را برطرف کنند .گليسريل تري نيترات مي تواند درد آنژين را سريعاً التبام بخشد . داروهايي هستند که مي توانند حملات آسم را سريعاً رفع نمايند و برعکس ، داروهايي هم هستند که بسيار دير نتيجه آنها اشکار مي شود ، به عنوان مثال در درمان با يک داروي ضد افسردگي هفته ها طول مي کشد تا شخص اثر مفيد آنرا احساس کند . اين امر منجر به اضطراب مي شود مگر آنکه پزشک از احتمال تأخير در بروز فوايد و اثرات سودمند دارو با بيمار صحبت کرده باشد .
* عوارض جانبي
عوارض جانبي شناخته شده دارو ممکن است در بيمار ظاهر شود . اين عوارض از واکنشهايي هستند که براي داروها پيش بيني مي شوند . دارو چه از راه دهان مصرف شود يا از راه تزريق و استنشاق ، در سرتا سر بدن توزيع مي گردد و اثرات آن محدود به يک بافت يا اندام خاص نيست .
به عنوان مثال داروهاي آنتي کولينرژيک که براي برطرف کردن اسپاسم ديواره روده تجويز مي گردند مي توانند بر چشم نيز تأثيرگذارده و موجب تاري ديد شوند ، بر دهان اثر گذارده و موجب خشکي آن شوند و بر مثانه اثر گذارده و موجب احتباس ادرار بشوند . اين گونه عوارض جانبي ممکن است بعد از عادت کردن بدن به دارو بتدريج ناپديد شوند ، اما اگر عوارض جانبي باقي بمانند بايد مقدار مصرف دارو را کاهش داد ، يا اينکه فاصله زماني بين مصرف مقدار دارو را افزايش داد.
عوارض جانبي برخي داروها مثل بعضي از داروهاي ضد سرطان مي توانند بسيار جدي باشند . اينگونه داروها فقط از اين جهت مصرف مي شوند که تنها داروهاي موجود براي معالجه يک بيماري خاص هستند بطوريکه اگر دارو مصرف نشود ممکن است آن بيماري کشنده باشد . اما همه داروها ، حتي ملايمترين آنها را بايد مواد شيميايي دانست که واکنشهاي جدي و سمي ايجاد مي کنند بويژه اگر نادرست مصرف شوند .
* مقدار مصرف و پاسخ دارو
همه در برابر يک دارو به يکسان پاسخ نمي دهند ودر بسياري موارد مقدار مصرف دارو بايد متناسب با سن ، وزن و سلامت کلي بيمار تنظيم گردد . مقدار مصرف هر دارو بايد آنقدر باشد که بتواند يک پاسخ مفيد بوجود آورد اما نه آنقدر زياد که منجر به بروز عوارض جانبي شديد گردد اگر مقدار مصرف دارو بيش از حد کم باشد ممکن است هيچگونه اثري ،چه مفيد چه نامطلوب ، بجا نگذارد . اگر مقدار دارو زياد باشد فايده خاصي ندارد و فقط عوارض جانبي بهمراه مي آورد . بنابراين هدف از درمان يا دارو آن است که غلظت دارو در خون يا بافت به حدي برسد که حداقل ميزان مؤثر و حداکثر غلظت مطمئن بوجود آيد به اين حالت طيف درماني مي گويند . طيف درماني براي برخي داروها مثل ديژيتال محدود است . لذا مرز سميت و اثر اندک مي باشد . داروهاي ديگري مثل پني سيلين طيف درماني بسيار گسترده تري دارد .
* واکنشهاي نامطلوب
واکنشهاي نامطلوب ، واکنشهايي هستند غيرمنتظره و پيش بيني نشده که به اثرات معمولي يک داروي عادي ارتباط ندارند . واکنشهاي پيش بيني نشده در اثر مصرف يک دارو مي تواند حالات آلرژي يا اختلال وراتثي ( مثل فقدان يک آنزيم که معمولاً دارو را غيرفعال مي کند ) باشد . واکنشهاي نامطلوب شايع از اين نوع عبارتند از جوش ، ورم صورت و يرقان که ممکن است بر اثر تداخل با داروهاي ديگر بوجود بيايند . واکنشهاي غيرقابل پيش بيني دارو معمولاً ايجاب مي کند که از ادامه مصرف آن دارو با نظر پزشک خودداري شود .
* اثرات مطلوب و نامطلوب
پزشک در ارزيابي نسبت ضرر به منفعت يک دارو بهنگام تجويز بايد فوايد و مضرات يک دارو را براي بيمار بسنجد . مثلاًعوارض جانبي چون تهوع ، سردرد و اسهال مي توانند از مصرف آنتي بيوتيک ناشي شوند . اما مطمئناً اگر عفونت خطرناک باشد . اين عوارض خطراتي قابل قبول هستند . اين عوارض براي قرصهاي ضد بارداري که چندين سال بايد توسط يک فرد سالم مصرف شود قابل قبول نيستند .
از آنجا که بعضي افراد نسبت به سايرين بيشتر در معرض واکنشهاي نامطلوب قرار دارند ( بويژه آنهايي که داراي سابقه آلرژي به دارو هستند ) پزشک بايد دقيقاً وضعيت بيمار و تناسب دارو را براي او مورد بررسي قرار دهد .
* پاسخ به شبه دارو يا دارونما
واژه « پلاسيبو » يا شبه دارو يا دارو نما به هر نوع ماده اي گفته ميشود که از نظر شيميايي خنثي است و به جاي داروي واقعي به بيمار داده مي شود . هرگونه فايده اي که از مصرف اين نوع دارو حاصل شود فقط به اين دليل است که بيمار معتقد است که اين دارو نتايج خوب بهمراه دارد و خود وي نمي داند که آنچه مصرف کرده است اصلاً دارو به معناي پزشکي نبوده است .
دارونما را طوري مي سازند که ظاهري شبيه يک داروي اصلي داشته باشد و به بيمار مي گويند که داروي اصلي را خورده است .
تداخل داروها
اگر دو داروي متفاوت را با هم بخوريد و يا دارو را با غذا ، مصرف نماييد ممکن است اثرات متفاوتي در مقايسه با هنگامي که يک دارو را به تنهايي مي خوريد در شما بوجود آورد . اين امر در بسياري از موارد مي تواند مفيد باشد و پزشکان براي بالا بردن تأثير درمان ممکن است ازاين ترکيبات استفاده کنند . غالب اوقات بيش از يک دارو براي درمان سرطان يا فشار خون تجويز مي شود .
اما تداخل هاي ناخواسته و زيانبار نيز وجود دارند . اين تداخل ها نه تنها بين داروهاي تجويزي ، بلکه بين داروهاي بدون نسخه نيز ممکن است بروز کنند . بنابراين بهتر است هشدارهاي روي برچسب دارو را بخوانيد و اگر دارويي را مصرف مي کنيد پزشک را در جريان بگذاريد . يک دارو با داروي ديگر يا غذا ، به دلايل متعددي تداخل مي کند .
* تغيير جذب
الکل و بعضي از داروها ( بويژه مخدرها ) فرايند هضم را در مرحله ورود محتويات معده به روده کند مي سازند . اين عمل باعث به تأخير افتادن جذب و اثر داروي ديگر که در همان زمان مصرف شده است ، مي شود . داروهاي ديگر به عنوان مثال ، متوکلوپراميد ( داروي ضد استفراغ ) مي توانند سرعت تخليه معده را افزايش داده و در عين حال جذب و اثر داروي ديگر را سرعت بخشند .
برخي از داروها با دارو يا غذاي ديگر در معده ترکيب مي شوند و ترکيبي بوجود مي آيد که به سادگي قابل جذب نيست . اين عمل مثلاً در صورتي رخ مي دهد که تتراسايکلين با قرصهاي آهن يا آنتي اسيدها مصرف شوند . شير نيز به همين روش جذب برخي از داروها را کاهش مي دهد .
* کاهش جذب در روده
جذب داروي A از ديواره روده در صورتي کاهش مي يابد که با داروي B ترکيب شود .
جذب يک دارو درصورتي کاهش مي يابدکه با مولکول غذا ، ترکيب شود .
* اثرات آنزيم
بعضي از داروها توليد آنزيمها را در کبد که کار تجزيه و شکستن داروها را بعهده دارند افزايش و بعضي ديگر نيز توليد آنزيمها را متوقف مي کنند يا کاهش مي دهند . لذا بر سرعت فعال کردن و غيرفعال کردن داروهاي ديگر اثر مي گذارند .
* دفع کليوي
دارو مي تواند توانايي کليه را در دفع داروي ديگر کاهش دهد و لذا مقدار دارو در خون بالا مي رود و اثر آن افزايش مي يابد .
* اثرات گيرنده
داروهايي که بر يک گيرنده اثر مي گذارند گاهي اوقات اثر تحريکي يکديگر را بر بدن دو برابر مي کنند يا ممکن است در اشغال محلهاي آن گيرنده با يکديگر رقابت کنند . به عنوان مثال ، نالوکسون گيرنده هاي مورد استفاده توسط مخدرها را بطور رقابتي مسدود مي کند و لذا کمک مي کند تا اثرات مسموميت مخدرها رفع شود .
چند مثال از تداخل هاي مهم
تداخلهاي نامطلوب بين داروها مي تواند از بين رفتن اثر مفيد يک دارو تا واکنش جدي بين دو داروي خطرناک باشد . برخي از واکنشهاي نامطلوب و خطرناک بين داروهاي زير رخ مي دهد :
داروهايي که سيستم عصبي مرکزي را خاموش مي کنند ( داروهاي خواب آور ، مخدرها ، آنتي هيستامين و الکل ) اثرات دو يا چند تا از اين داروها به صورت همراه مي توانند اعتياد آور باشند و يبوست خطرناک پديد آورند .
داروهايي که قند خون را پايين مي آورند مثل سولفوناميدها و الکل ، اثر داروهاي کاهنده قند خون را افزايش داده و لذا قند خون را بيشتر پايين مي آورند .
ضد انعقادهاي خوراکي و داروهاي ديگر بويژه آسپيرين و آنتي بيوتيک ، با هم تداخل ايجاد مي کنند ، چون که اين داروها مي توانند تمايل به خونريزي را افزايش دهند پس لازم است اثرات آنها در هر مورد بررسي شوند .
بسياري از داروها و غذاها مي توانند باعث افزايش شديد فشارخون ، هنگام مصرف با اين داروها شوند . از آن جمله مي توان به آمفتامين ها و ضد احتقانها ، غذاهايي مثل پنير و شکلات اشاره کرد.


 آمار
وب سایت:
آمار
وب سایت: